


Shark Olfaction, Electroreception, and Quantum Coherence within the Synapse

(Link to Audio recording narrated by Author)
After countless hours of deliberation, I have decided that I want to share an idea that has enraptured the entirety of my life for longer than I can remember. This will be the first time that I have ever coherently elucidated this idea even to myself, nevermind to the world. I had originally hoped to establish this as a plausible direction to my thesis for my potential graduate/doctorate career. I now hope that somebody else may find interest and pick up the duty to unravel these mysteries.
This means MORE than everything to me. In a very real sense, this paper may be the most comprehensive existing description of my life's most meaningful familiarities, and it is because of that importance that I recognize it is absolutely imperative for me to outline the background in terms of its contexts. Bear with me as you read the beginnings of this college-essay-esque exposition of my background.
For those of you who have known me since I was a child, the first thing that comes to mind for most is my obsession with sharks. I have been dubbed “shark boy” for longer than I can remember. My most rooted and core memories are of my childhood bookshelves filled with hundreds of shark books (that I still own) that I had scoured passionately over ever since I learned to read. In fact, I vividly remember gaining the desire to learn how to read for the sole purpose of being able to learn about sharks. It was in this era that I gained my first memory of UCSD as well, inspiring me to become a marine biologist one day.
It was sharks that got me into science and even cars, after admiring the mesmerizingly hydrodynamic design of these aquatic sharp-toothed missiles. In fact, it was from the research uncovered on the hydrodynamic boost provided by the dermal denticles on shark skin that revolutionized (and sparked a controversy) within the competitive swimming scene, which promptly inspired me to take up competitive swimming for a brief era. From then on, I slowly lost interest in sharks due to the absence of any novel ideas or research. It was then that I moved onto venomous reptiles, where it became clear to me that my unharbored desire was directing me into the molecular and biochemical realm.
It was then that I realized that I had hit a roadblock. No matter where I had turned, all of the molecular and biochemical profundities I discovered lead me to the same giant monolith of impossible quantum coherence in complex physical systems like biology. This mystery did not go away, and in fact worsened in confusion and anomalous complexities the more I studied it. It can be said that in a similar manner to how mathematicians/theoretical physicists bang their fists on the table when they accidentally break causality, biochemists bang their fists on the table in frustration when they encounter quantum behavior (read: physics envy).
Moving onwards, I went to consecrate my efforts into understanding the strange quantum behaviors that lay at the bottom of neurobiological principles like consciousness and synaptic/neuro plasticity. It was during a particular meeting with a Professor that I brought up the notion of surmising that perhaps the particularities of cognitive memory (which cannot simply be chalked up to “hippocampal activity”) were very intimately correlated with the various patterns and proportions of extracellular matrix proteins expressed on the post-synaptic terminal. I continued by propagating the logic towards the understanding that it must be the MAGUK proteins responsible for the regulation of the various key scaffold proteins like SAP-102 and post-synaptic densities like PSD-95 (or even pre-synaptic markers like Bassoon) which regulate and provide anchorage for the ECM proteins, providing a feasibly shallow explanation for how quantum mechanically coherent signal transduction could be manipulated and specified via the electrically active residues of ECM proteins in the vicinity of the bound receptor domain. This provides synaptic transmission with an extra degree of freedom for a total of two in input: the neurotransmitter species, and the electron-binding-dependent voltage. This, combined with the suprathreshold/subthreshold integration mechanisms accumulated over a neuronal junction patch containing a large quantity of synapses, seems plausible in that it generates a larger modality for manipulation and increases the number of possible modular output computations. It was then that Professor Zou humbly informed me that this was actually the personal theory of the late Dr. Roger Tsien, UCSD Nobel laureate for the discovery and innovation of GFP (Zou, the Lab I would later go onto volunteer in). I went on to read (parts of, the mathematics defeats me) Roger Penrose’s The Emperor’s New Mind and Shadows of the Mind. In the former, I was sufficiently convinced of Penrose’s deduction in regards to the non-computational nature of consciousness, which made appreciable sense considering what the current science has revealed about the dynamics of signal transduction at the receptor level. In the latter, Penrose along with anesthesiologist Stuart Hameroff, extends his theory by considering the quantum coherence properties demonstrated by the microtubulin structures within the axonal terminals that heed the biochemical facilities necessary for proper synaptic transmission (Edit: 2024, Quantum-Coherence within wet-warm microtubulin structures of Biology gains yet more evidence). I saw this as yet another key piece of evidence for Tsien’s theory, as it has been revealed today that the dynamics of synaptic microtubules are regulated by an unintelligible mixture of MAGUK proteins, the PCP (planar cell polarity) pathway, and potentially more. Nothing irks a Biochemist more than discovering that their pathway is ultimately regulated by G-protein coupled receptors, Prof. Susan Taylor’s PKA, Ca2+ influx, or some other macrospective phosphorylation cascade, because such is to say that the regulation dynamics of their model will be existentially impossible to compute entirely. Consider Dr Sydney Brenner’s golden child of C. elegans. The 1000 celled nematode now has a complete cell fate map and a complete connectomics adjacency matrix, and yet even so we are unable to intelligibly decipher conclusively the chemical dynamics of living activity. The complexity that evolves from the accumulation of each quantum mechanically probabilistic event seems to appear superficially deterministic in these gigantic, humanly incomprehensible regulation networks. As Sir Isaac Newton once said, “If I see far, it is only because I am standing on the shoulders of giants”. At this point I had already most exquisitely and deeply fallen in love with “Quantum Biology” as a field, with the most peculiar examples to me being: the coherence/decoherence mystery at the synapse, and pretty much all of olfaction, if we are even at all intended to distinguish between the two as separate examples.
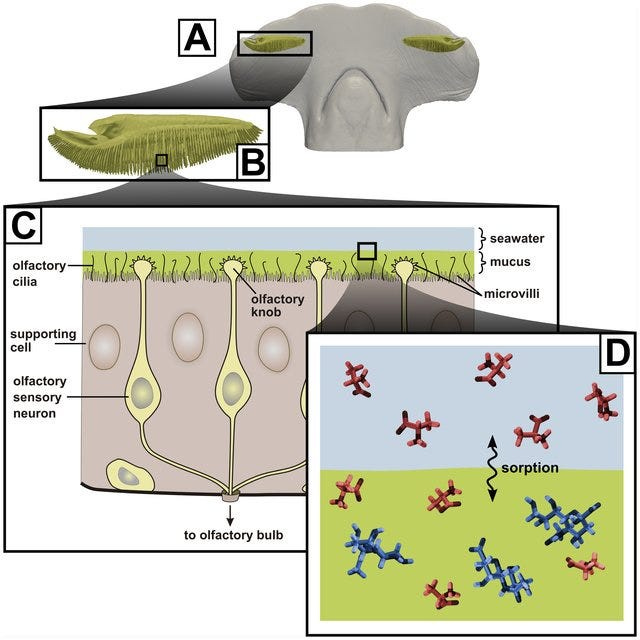
If you aren’t familiar, the olfactory capabilities of every organism is not understood at all. It is absurdly impossible to conceptualize how it even makes sense that we can detect odors that exist in the tiny quantum mechanically coherent concentrations that they exist in. The idea here is coherent delocalization, which suggests that the membrane-bound receptors themselves exist in a state of quantum coherence, allowing them to simultaneously sample and bind multiple probabilistic molecular conformations at once. I implore you to sit down for just a moment and attempt to comprehend how it is possible that tiny molecular receptors in my nose and nervous system can even begin to interact with the miniscule concentration of odor molecules diffused all throughout the air. For example: Humans are odor sensitive to the molecule geosmin (contributive to the smell of rain, petrichor) in concentrations as little as 5 parts per trillion. Assuming a perfectly classical reign of Physics onto Biology renders no explanation for this, it implies a quantum mechanical realism into Biology that suggests coherence persists through “wet & warm” systems somehow despite the obvious challenges that entails.
Edit (August 2024):
It’s been almost more than a year since I wrote this essay. I have since learned much about this topic since then, which I feel some itchy obligation to re-insert into here post-factually. This lengthy insert — wherein I go into much deeper depth of the topic — comes from another pseudo-essay of sorts that I am working on (something I’d like to call my pseudo-biographical-résumé). Forgive me if it falls outside of context or seems a bit misdirected on tangents.
Enter Quantum Biology, where all of this led me to taking one of my favorite classes yet in hopes of understanding a bit more of the quantitative-stochastic roots of Biological processes. I learned much from said class– much of which I have already described here– but really, as always, I came out with more questions unanswered than when I had began. As such, my quantum frustrations with biology only grew but in at least irritably meaningful to get me to start taking my PhysMatics explorations more seriously and rigorously.
“When a distinguished scientist says something is possible, he’s most likely right. When such a scientist says something is impossible, he is probably wrong”
-Arthur C. Clarke’s 1st law
The common critique against quantum phenomena in Biological setting is that “it is too wet & warm of an environment to possibly preserve coherence” as heuristicized with the balls’in’a’box model: large systems with noisy energy fluctuations lose their information into the dissipative heat bath of the surroundings, the system loses its phase coupling with itself and becomes coupled to the environment. They effectively become lost from another in coupling with the mass interaction with the environment– much the way a few raindrops in a pool retain their identity of which ripples are engagedly interfering, but when it has escalated into a rainstorm, the surface of the water becomes an entangled mess– hence the term, “decoherence”.
And yet we see the unassailed observation of quantum phenomena prevailing in Biological systems nonetheless: electron tunneling in Photosynthesis’ OEC, enzyme catalysis, and cytochrome c. The prevalence of magnetoreception in birds utilizing a spin-sensitive cryptochrome to detect the earth’s weak geomagnetic field despite the world’s noisy E&M environment (effectively a biological analogue of the stern-gerlach experiment). And my favorite case phenomena, olfaction, the sense of smell. For example: Humans are odor sensitive to the molecule geosmin (contributive to the smell of rain, petrichor) in concentrations as little as 5 parts per trillion. Assuming a perfectly classical reign of Physics onto Biology renders no explanation for this, it implies a quantum mechanical realism into Biology that suggests coherence persists through “wet & warm” systems somehow despite the obvious challenges that entails. Here at last the circling dorsal fin breaks the surface in becoming manifestly apparent of how sharks fit into all of this.
As with the example of human proclivity for petrichor, it’s not that Sharks in particular that behold a mysterious mechanism of olfaction– albeit they do have a canonically strong sense of smell (especially in consideration of the electrochemical noise of the ocean), but so do many other animals– but it is in the conjunction of their other sensory abilities which interests me so. Namely that sharks possess other adjacently mysterious sensory capabilities like electroreception and magnetoreception, the latter of which stands to some level of debate of whether they are explicitly detecting magnetic fields or if it is just some derivation of electroreception (since E fields and B fields share Maxwellian progenitor). All three of these sensory faculties– olfaction, electroreception, and magnetoreception– share a Physical inquiry which unites them, they are fundamentally an exhibition of the decoherence mystery. That is, heuristically: how quantum information can persist in preserving coherence to be detected by Biological systems despite the wet, warm, noisy environment and all it entails. One may speculate therefore that these three sensory faculties are actually underlied by the same mechanistic profundity. Looking at the morphology of these infrastructures may give us some further credence to their closely-related overlappings.


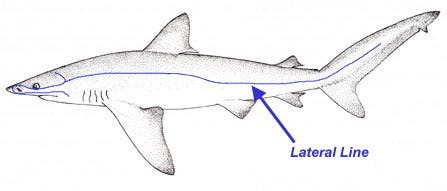
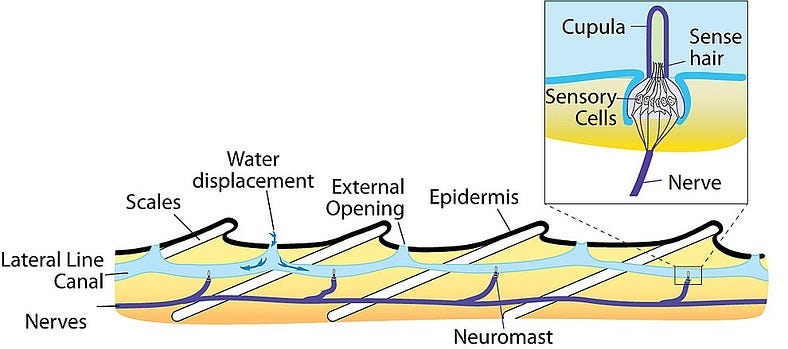
Electroreception) mediated by the “Ampullae of Lorenzini”, which are these small pores on the snout of a shark immersed in a glycoprotein jelly substance prowessed with semiconductor-like properties. These sacs are lined to afferent neurons, acting as electroreceptors via voltage-dependent calcium channels triggering depolarization in these unique ribbon synapses, also equipped with a peculiarly expressed potassium channel displaying anomalously fast repolarization kinetics allowing for unexhausted quick repeat firings without diminished conductance despite substantial abuse of synaptic activation/deactivation (a great clue here in understanding the mechanism of what Kalmijn alluded to above in terms of how signal may be amplified via positive feedback loops to rise above noise, as well as the Quantum Zeno effect which I will get to later). Interestingly, this biological infrastructure for electroreception seems to be the evolutionary product of a more basic mechanoreceptive form which allow sharks to detect harmonic disturbances in the water–say of a struggling injured fish– via their lateral line, employing a similar structure of pores equipped with sensitive hairs which relay signals to the afferent neuron dependent on the sway of water displacement as it enters the canal. There is also the question here of how the shark ought to tackle noise, vibrations of the water column produced by its own movements, where it has been found that an inhibitory efference copy of cholinergic signals (corollary discharge) are matched by the brain to its according muscle movements which nullify the detection of its own movements. There is more to be said about electroreception but I sense that it may come more lucidly if I leave it to be dissolved in more context for later.

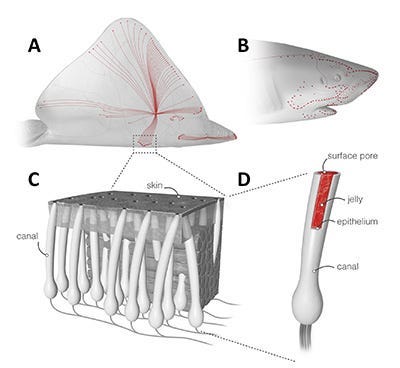
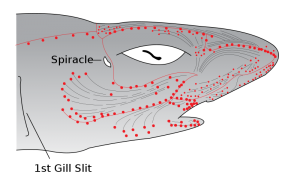
Magnetoreception) The question of a magnetoreception sensory capability in sharks (and other electrosensory organisms) is quite a convolutedly difficult idea to parse exactly in distinction from its electrosensory capabilities, considering all of the Physical kinship entailed by the two phenomena. I fear that to explain the full extent of this relationship would require something akin to a couple semesters of an electromagnetism course, and as such please forgive me for blasphemous heuristics I will inflict here to Maxwell’s equations. Magnetic fields are produced by moving electric charges, and similarly electric fields can be generated simply by a changing magnetic field. Thus, one can imagine it as such like a charged particle bobbing up and down on the surface of an ocean, whereby the particle produces waves by means of its movement but also recursively rides its own wave which it produces. The particle produces waves by bobbing, and by riding this wave it causes the particle to further bob up and down, thus producing more waves, etc. This is actually the nature of light, hence why it is called an electromagnetic wave.

Further diving into the physics of this phenomena, we have to define what exactly we mean by “a moving charge”. This requires the installation of a relativistic notion, structured by what one might picture as reference frames, perhaps elucidated most famously by Einstein’s thought experiment of weightlessness in a falling elevator. Allow yourself the conceptual entertainment to digest this: a stationary observer watches as a charged particle moves left to right in front of them, producing a magnetic field. This is Physically equivalent to if the charged particle were stationary, and the observer were walking right to left of it, where they would see it produce magnetic fields of the same kind. This is an essential structure of the relativistic implications of electromagnetism, that the charged particle itself need not actually be “moving”, it needs only to be moving in reference to something else, in this case the observer. One can imagine that placing a perfectly stationary charged particle on a table, and simply by the virtue of walking around that table one would observe a magnetic field, since the “movement” of the particle is indeed happening for its personal reference frame.


Let us presume for a moment that Sharks are honing in on what appear as magnetoreceptive capabilities purely as a spandrel by means of its electroreceptive infrastructure. It is perfectly fitting to export this concept of fields generated by motional charges to the ocean, whereby one can imagine ions carried by the water currents are a source of electromagnetic fields. And even further, by bestowing the frame of reference not on the moving body of water but on the shark-observer themselves: “Alternatively, a directional compass sense may be facilitated by a comparison of inputs from the vestibular and electrosensory systems, making use of directional asymmetry in the voltage drop across electroreceptors during movement of the head28. That is, as the animal swims, the head moves side to side in a sinusoidal manner, producing phasic/cyclical stimuli to sensory maculae within the vestibular system, as well as a voltage drop across the ampullae of Lorenzini corresponding to the change in head position relative to the external electrical field as the animal moves (Anderson, Clegg 2017)”.

That seems plausible, and perhaps in some sense a conjunctive truth not subject to the law of the excluded middle (i.e: Sharks use both their electrosensory and some separate magnetosensory infrastructure in yielding their capabilities to detect magnetic fields). Although– not to get in on the controversy as it is besides the underlying point– there is also some evidence which would suggest otherwise:
“Reversible sensory impairment methods were used to test the hypothesis that receptors for the perception of magnetic stimuli may be located in the naso-olfactory capsules of sharks. This hypothesis invokes the existence of magnetosensory structures within the olfactory organs, perhaps homologous to systems described in some teleost and avian species23, 46, 55 … Given these experimental conditions, we attempt to narrow the possibilities regarding which sensory pathways are involved in magnetic field perception, and whether magnetic fields per se (rather than an electrical derivative) are detectable by sharks (Anderson, Clegg 2017)”.
There is some considerable overlap here between shark olfaction, magnetoreception, and electroreception looking purely from the standpoint of morphology. In fact, naively it is often mistaken that the ampullae of lorenzini (considering their position and proximity to the nostrils) are responsible for their sense of smell, and there is quite a narrow margin in distinguishing that shark olfaction actually comes from their “nostrils” which are considered unrelated to their ampullae also localized on the snout. This magnetosensory infrastructure is believed to be colocalized with the olfactory organs, further blurring the lines and giving reason to suspect that all three of these senses are in some sense deeply related (which they are, at least on the basis of the physical principles which underlie them). But for the moment just as a means to clarify magnetosensory mechanism in particular, let us move away from this particularly blurry case of sharks and look at the case of another organism who is equipped only with magnetoreception as to isolate it into focus not as an epiphenomena.
Let’s look at the mechanism of magnetoreception in avian species, as most are colloquially familiar with their navigational prowess by way of the weak geo-magnetic field. Hopefully as to not get too deep into biochemical/physical technicalities, I ask that you forgive some of the rather injective uncontextualized concepts I will mention, and that I ask you may have to look some things up on your own. It starts with the premise of wondering how such a thing like the earth’s weak geomagnetic field (amidst all of the noise)– magnetism effectively being a macrospective property of quantum spin (a bit of a heuristic)– can possibly be detected by organisms if taken under the assumption that biological systems are too wet & warm to retain the coherence of quantum properties. The mechanism predicates on a particular class of photosensitive flavoproteins called cryptochromes, most commonly known for their function in circadian rhythm regulation, and it is exactly this photochemical dependency which underlies its action.
It is called the radical-pair mechanism, whereby a photon striking this cryptochrome allows for photoexcitation creating an entangled “radical pair” of electrons, that is two lone electrons (called radicals) which each have a unique spin state entangled to one another. These electron radicals can either have aligned or opposite spins (a heuristic of the singlet 𝑆₀ vs triplet 𝑇₀ states), sensitively dependent on the angular orientation to an external (in this case, geo-magnetic) magnetic field. “The state of these spins then evolves under the combined effect of the Earth’s weak magnetic field and internal nuclear hyperfine interactions with the host nuclei. Finally, the rate of charge recombination depends on the spin of the separated charges, directly influencing the reaction products of these radical pairs. These differing reaction products are in principle biologically detectable. Thus, if the relative weights of the singlet and triplet states are sensitive to the angle of the external (geo-magnetic) field, the reaction products will be also, leading to a magnetic compass […] The 𝑆₀ and 𝑇₀ states are equivalent to Bell states, maximally entangled states that are highly desired in quantum information schemes […] Essentially, some dephasing models leave the sensitivity of the radical-pair reactions to the external fields mostly intact […] In addition, because these disrupting fields were so weak, of the order of 50–100 nT, they demand that the radical-pair model maintain quantum coherence of its spins for tens of microseconds. It seems then that this disruption due to external fields is a purely quantum effect, and that both the quantum coherence and entanglement properties must be sustained for timescales ‘exceeding the best man-made molecular systems” (Lambert, Chen 2012)
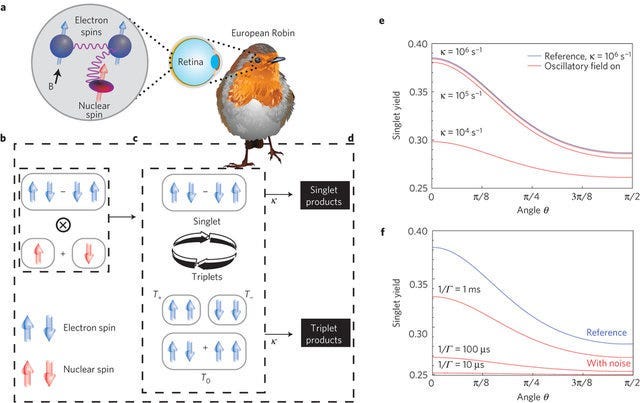
This of course sounds extravagantly impressive for Nature to have engineered at the biological level, hence the skepticism of the mechanism and proponents of alternative suggestions identifying iron magnetite deposits in the beak as the biological compass (which have been shown actually to be macrophages and unrelated to magnetoreception). Although when taken into biophysical generality, it is not all that outlandish considering that the underlying mechanism of photoexcitation-related quantum engineering is prominently conserved throughout Nature’s organisms. Some cases exceptionally homologous to avian cryptochromes include the FMO trimer complex in photosynthesis’ reaction center, and the OEC and P680 excitation in photosystem II.
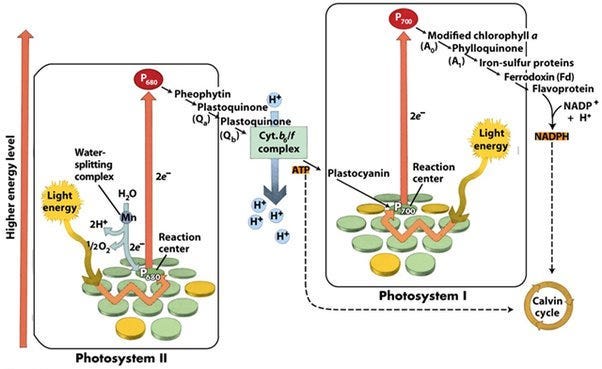

Hopefully not to sidetrack with too much detail, but the light-harvesting complex of photosynthesis as a whole ought be taken with a sharp eye in recognizing that it necessarily requires the employment of both classical and quantum phenomena in giving the near 100% energy efficiency yield that it is observed to. Taking a purely classical purview makes such a process seemingly impossible, as it necessitates photons must reach their reaction sites before the energy dissipates on a sub-nanosecond time scale. Somehow this process (particularly the FMO complex, which is mediating this energy transfer between chlorosomes and reaction center) is able to leverage quantum coherence allowing chlorophyll molecules to enter an excitonic state sharing the energy of the photon across different molecules. The existence of these excitonic states allows for coherent energy transfer — meaning that the energy can “explore” multiple paths simultaneously, enabling it to find the most efficient route to the reaction center. The literature evidence has shown that the FMO complex is capable of sustaining considerably remarkable periods of coherence, permitting the energy of the photon to exist in a superposition of states, traveling all possible routes within the antenna complex at the same time. When this photon finds the correct destination, the superposition collapses, offering available energy. There are more peculiarities to this story, but again I fear I may draw too long of a tangent. For example, the FMO complex is in fact quite antithetical to the “wet & warm” dissidence, as rather interestingly it seems capable of in fact harnessing what’s called “noise assisted transport” in aiding this hopping of energy states, weaponizing what would be considered a destructive element to quantum coherence as an effective aid to its function.

All of these biophysical case studies may present themselves somewhat hazily, and perhaps it is by drawing their uncannily representative analogues to Physics-proper experiments in which they become more fleshed out in apparent lucidity. Just starting with the more obvious one whose analogy requires minimal context, one can see that the quantized spin dependent radical-pair mechanism of avian magnetoreception is most analogous to the Stern-Gerlach experiment. Whereby a beam of silver atoms are sent through a chamber enforced by an inhomogeneous magnetic field, flinging the atoms according to their spin onto some detection screen and showing a discrete distribution. It’s one of my favorite demonstrations of the fundamental principle of quantum theory (superior to Young’s canonical double-slit in my opinion) as it brings clarity to the nomenclature with which the word “quantum” is defined. Just as it’s easy to understand the discretized nature of magnetic poles, it shows that spin at the quantum level is also discrete seeing as to how the experiment does not produce a continuous distribution as one would expect if angular momentum were to be seen classically as being a continuous value. One can also understand the basic heuristic of what decoherence may represent in this context: that atoms at the individual small-scale (quantum) strata exhibit their discretized nature of spin (up/down, either one or the other), whereas a large macro chunk of silver will exhibit something like a measured continuum being comprised of a large mixture of atoms (say something like, 78.9% spin up and 21.1% spin down) where the “discreteness” of each individual atom is summed and melded away into collective blurriness.

The case of shark electroreception, or even its adjacents like the electrostatic forces utilized by plants and pollinators as navigational tools, speak too of a profound Physics experiment. The Bohm-Aharonov experiment done in the late 1960s, is one of my favorites for several reasons. It touches upon the essence of many philosophical pivots which strike my fascinations most dearly:
1) We don’t know what we don’t know. There’s a great adage along the lines of “Even Einstein carried his luggage”, an expression of the fact that something so trivial like the modern rolling luggage was discovered/invented only in 1987, something seemingly so obvious and powerful yet elusive of the greatest minds for centuries past. The same can be said of electromagnetism, largely considered all wrapped up and done by that time, none the wiser that this experiment would show reveal that the true underlying profundity of electromagnetism had yet to be understood even so late into the game of the 1960s (think of Planck’s advisor Philipp von Jolly telling him that it was probably not a great idea to study theoretical physics, since there was not much left to do).
2) What, and Where do the demarcations and conjectives truly lie between the continents of Mathematics and Physics? It’s a topic I’ve amused expeditiously throughout many of my pseudo-essays, and in particular this serves as quite a cogent example of the not so trivial distinctions. There are often times in which something which we think to be of real Physical substance turns out merely to be a fictional mathematical aid. There are also times in which we think something to be purely auxiliary mathematical gadgetry, but turns out to be a real Physical quantity. Examples include: Dirac’s negative energy solutions– thought to be mathematical chaff in the same way one typically considers only the non-negative solutions of the √ operator– turning out to be legitimately Physical antiparticles (or perhaps controversially even the ghost/virtual particles haunting the standard model Lagrangian, are they merely mathematical deus-ex-machina’s or representative of real Physical objects which we have yet to find means to observe?). The Bohm-Aharonov experiment is specimen of the latter example, whereby it was previously thought that the Efields and Bfields were the real objects of Physical primacy and that the Gauge potentials were just tools of Mathematical redundancy. It wasn’t until this experiment that we understood it was actually the potentials which were the true underlying substance of electromagnetism– especially when put under quantization– and that the Efields and Bfields are really just something like derivative epiphenomena. The experiment shows this rather beautifully, by shooting charged particles around a perfectly insulated solenoid (externally, Efields and Bfields = 0) and showing interference effects due to the fact that they are actually coupled to the potentials. It is uncannily provocative to demonstrate that this experimental setup is capable of detecting whether or not a current is being run through a perfectly insulated solenoid, that despite the absence of any explicit external electric/magnetic fields, one is revealed to the fact that it is really the intangible fabric of scalar and vector potentials which are the star of the show.


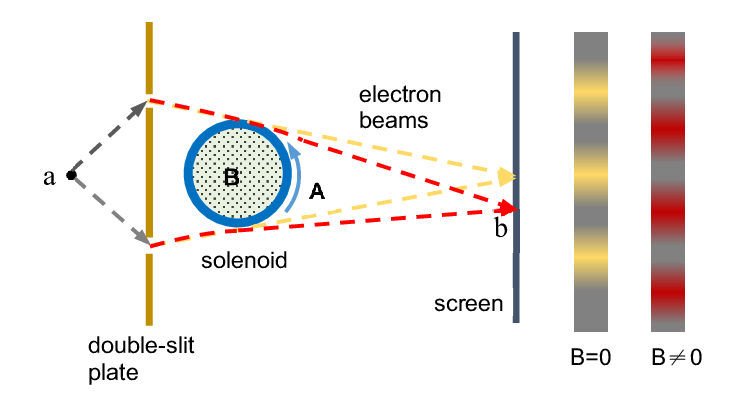
As C. N Yang put it: “[he] considered the AB effect to be only direct experimental proof of the gauge principle. The philosophical import is that the magnetic four-potential (ϕ,A) overdescribes the physics, in that all observable phenomena are the same after we perform a gauge transformation. The Maxwell fields underdescribe the physics, as it does not predict the AB effect. And just as the gauge principle predicts, the quantities that are invariant under gauge transforms are exactly the physically observable phenomena.[10][11]”. Gauge invariance, in hindsight turned out to be an enormously fruitful line of insight in the world of PhysMatics (As Yang knew very well in formulating the Yang-Mills-Maxwell equations generalizing electromagnetism in the language of a gauge group). It is essentially the discovery that, what appeared to be previously simply a mathematical redundancy from the properties of calculus (the way that a +C constant disappears under derivation), actually turns out to be a new landscape of freedom on which to play on. It says that the different choices of potential functions can still yield the same resultant electric and magnetic fields, and that in fact this new topological buffet allows you to, as Juan Maldacena put it (although really taken from Pia Malaney’s PhD thesis with Eric Weinstein): find that the laws of Physics ought to obey exactly in the same way irregardless of your choice of “currency rate”, in the context of choosing d/dx = 0 as your “reference level”. Again, there’s much more to be said about this but I fear I am already tiptoeing on a tangent out of the scope of relevancy.
In one of my pseudo-essay pieces I take it into quite the poetically generalized thema, that “the potential is more primary than the substance”. Whether that’s in geopolitics ala Otto von Bismarck’s “politics is the art of the potential, the attainable…” in terms of strategic possibilities and potential threats being more primary than the actions themselves. Or in the predictive nature of neurobiology– thanks to the lag of axonal conduction that our sensorimotor capabilities must account for– that we almost aren’t ever truly reacting to catching a ball but instead we are predicting it. Or even when it comes to the stacked tranches of financial derivatives (very aptly named) where you have more money riding on the bets of some asset rather than the underlying asset itself (say synthetic sOIL futures versus the raw asset of petroleum itself).


The point being here that, as beautifully shown with the Bohm-Aharonov experiment, perhaps the underlying prowess of electroreception is not so much to do with the weakness and noisy imperceptibility of explicit Efields and Bfields but rather– fitting into a quantized narrative– to do with some capability of coupling to the scalar and vector potentials. Such that instead of banging our heads against the wall in questioning how signal can be obtained from such negligible Efields and Bfields, perhaps the answer lies in understanding those fields merely as epiphenomena and recognizing instead that the underlying object of potential grants much more liberty of consequence as a non-local influence. Incurring the technical debt of postulating some sort of mechanism by which biophysical electroreception couples to the potential fields allows you to bypass the main irredeemabilities of looking at it purely from a classical ledge. Sure, now you are beholden to explain some sort of quantum biological mechanism in explaining the infrastructure of electroreception, but with this investment you at least eliminate certain pivots that being the question of the Efield’s decay over distance (as per the inverse square law). After all, this is the check-kiting nature of the theoretical pursuits, often requiring you to take some assumptional advance as investment debt to patch some surface level inadequacies. It’s why theoretics is often considered a whack-a-mole endeavor or an intellectual hydra, whereby with every meaningful solution you incur some greater debt elsewhere that renders potentially bigger questions to quench.
Anyhow at the end of the day, the ineluctable red string of this quantum biological cobweb is the question: in all of these cases of context, whether it’s a pigeon stern-gerlach experiment or a sharky coupling to the electromagnetic 4-potential, how does Biology preserve the ability to habituate these quantum properties in the obvious face of challenges that is the noisy, wet, warm environment that it entails? In my mind there are four possible strategies so far as I can think of (some of which I deem more plausible than others, which I hope is self-evident by their weight of their descriptions):
1) Structural protection.
One might imagine, say in the context of avian magnetoreception that the protein structure of the cryptochromes themselves (or perhaps just some particular subunits, or the surrounding proteins in which they are enclosed) are imbued with some kind of faraday cage-esque properties that protect the radical mechanism’s microenvironment from noise. I hesitate to say much more in that I can’t exactly be sure of the details of how such a thing could be done. Although, in some sense you may consider this an offshoot or even a subset of bulletpoint 3) to follow.
2) Speed.
As has been shown in the case of photosynthesis’ PSI/II (P680 and the likes), the dependent reactions (charge separation, and excited state formation) happen on the scale of nanoseconds to picoseconds. As Zewail’s 1999 chemistry Nobel prize on femtosecond laser spectroscopy– and further, the recent 2023 Nobel prize on attosecond spectroscopy– have revealed: much of quantum spookiness in regards to the transition from coherent states to decoherent states are really just phenomena harbored in the domain of ultra fast physics. As temporal resolution advances show that the femtosecond scale define photon and electron interactions, and that the attosecond scale allows one to peer even deeper into electron dynamics (like showing that orbital transitions actually happen at variable speeds and not instantaneously as once thought), it may be fair to say that one of the most characterizing distinctions between classical and quantum reigns, is not simply size but the time strata of which their interactions hold most significance i.e: that the femto↔attosecond frontier is where quantum effects dominate. If we are to suggest that the blurry transition between classical and quantum is, in some part, dependent on the strata of time in which the observation is being measured, then one can adjoin the deduction that in order for Biological systems to make use of such quantum behaviors that it does so on the basis of high-speed dependent reactions. For if you are capable to operate on such small temporal windows, you can beat the rate at which decoherence occurs– as phrased in the context of photosynthesis radical states: The procedure of adiabatic elimination46 can be applied to quantum systems with dynamics occurring on widely separated timescales. The effective dynamics for a ‘slow’ system of interest are derived by assuming that a ‘fast’ system process is completed on a timescale where the ‘slow’ system is static (Marais, Sinayskiy 2015). That is, put theatrically, “you put pedal to the metal not to go fast, but only so that the other cars stay still”. We know certainly that this is the case at least in the electronic and photonic interactions of photosynthesis, with various studies showing much of the activity in the reaction center (e.g: formation of delocalized excited states, charge separation, charge transfer, etc) happening in the femto-pico second time scale. Even better, there was recently a paper that was able to map the electronic decoherence pathways of thymine in water via rudimentary Raman spectroscopy, occurring roughly on an interval of ~30 femtoseconds. Further, they were able to modulate part of the molecular structure via chemical substitution– in particular, the ring– and control the rate of decoherence as dependent on the fidelity of hydrogen-bonds formed with the water solvent. The paper seeks to understand the molecular rationale behind its connection to decoherence rates in an effort to modulate it, much the way enzymes have naturally done so in optimizing their binding sites (enzymes quite literally being the gold-standard of quantum chemistry, if we were able to produce synthetic enzymes with their binding & reaction efficiency thanks to their optimization for tunneling and whatnot: it would pretty much revolutionize the entire industrial workflow regarding catalysis, and likely break the way for new highly generative sources of energy- say a synthetic photosynthesis replicating or even surpassing the near 100% e=hv yield it already exhibits), but for the purposes of this paragraph I just mentioned it as a note for the ~30 femtosecond emphasis. Keep all of this in mind as it may give warm relevance to bulletpoint 4).
3) Quantum error correction/Topological trickery.
As with the avian magnetoreception radical pair mechanism which depends intimately on spin, one can’t help but entertain the plausibility of error correction techniques as have been thoroughly utilized in the field of quantum computing as well as some other preservatory techniques in topological quantum computing. It’s definitely going to take a little more than my plate can fill in order to explain the intricacies of how error correcting algorithm works– but essentially it is like using an algebraic expression to preserve information, for example with some sequence of numbers: 2, 4, x, 9 where x is the missing bit of information that has been lost for whatever reason, one can mitigate the risk of losing information by simply adding one more entry 2, 4, x, 9, 20 where the fifth entry is simply a sum of the first four, whereby one can deduce that the missing bit x = 5. This system relies on redundancy, in that you must add this extra piece of information– the sum– which already contains the patterned structure of the data it seeks to protect (as you can quantify with Shannon’s entropy, which I shan’t elaborate on but you may like to read on in a pseudo-essay I wrote) You can do something of a similar (but obviously much more complex) nature with quantum computing whereby you smear information about one qubit over the n other entangled qubits. In the biophysical context, one might postulate that this robustness against decoherence may exist as a spin-orbit coupling mechanism (Ferritin and metalloproteins?). The concept relies on magnetic anisotropy (much like what underlies NMR spectroscopy, which I wrote my IB HL Chemistry IA on back in highschool. A good chunk of my first enamoration with quantum weirdness) whereby you can consider that the electron not only has a spin, but also a felt magnetic field from its frame of reference as it orbits around the nucleus. As what’s called “electron shielding”, the spin of the electron will seek to interact with the felt magnetic field by way of Ampere’s right hand rule i.e: if the magnetic field points upwards, then a counterclockwise spin would counteract the felt magnetic field as it generates a downward magnetic force. This coupling, between the orbit of the electron and its intrinsic spin, can help stabilize quantum states as it effectively locks together its intrinsic spin and the orbital angular momentum, making it more robust against decoherence. Another similarly plausible avenue by which Biology may have developed as an error corrective/coherence preservation technique is utilizing the nonlocality of topological information, especially prevalent in the corner/edge states of self-repeating fractal structures. I wrote more about this here if you’d like to read it in greater detail, but essentially the premise is that– as recently shown in a paper on using bismuth fractal crystals as a substrate– topological properties are invariant throughout a structure, regardless of whether you are looking at just a corner or the structure as a whole (thanks to its self-repeating nature), and thus you can use the these corner states as qubits given that their topological properties are conserved nonlocally. The fact that these corner states are spatially isolated protects them from local noise and decouples them from bulk state perturbations, providing them robustness as the existence of their properties is guaranteed by the topology of the structure as a whole rather than their local properties. One might proceed to conjecture that the fractal motif which appears ubiquitously throughout Biological structure is not so much a pursuit of energetic gains (as was found in the case of citrate synthase’s fractal structure), but rather that its purpose is of an informatic nature– say in preserving quantum coherence.
4) The Quantum Zeno effect.
This is a rather counter-intuitive notion, that in some quantum systems, frequent measurement or interaction with the environment can paradoxically slow down decoherence, a phenomenon known as the Quantum Zeno Effect. In other words, as aforementioned in 2), spamming measurements/interactions of a system with such high interval frequency in fact slows down or even halts its evolution, thus the associated sentiment “a system cannot change while you are watching it”. Its moniker comes inherited from ancient Greek philosopher Zeno’s arrow paradox, which posits similarly that because no movement is seen during any single instant slice of an arrow’s flight, it cannot possibly be moving at all. Understandably this sounds very hand wavy– which is quite alright, I did not intend for this to be a textbook– but just to enunciate a bit more conceptual clarity perhaps allow me to go into a bit more technicality which I must ask for your forgiveness if there are any lapses in rigor. The counterintuitivity here is that while we know measurement seeks to collapse the wave function 𝜓, describing a superposition of states, into one particular eigenstate of the measurement basis, the point that the Zeno effect attempts to captivate is in inhibiting the system’s time evolution away from the initial state (as the Schrödinger equation tells us). Because quantum evolution state transitions require time, by spamming measurements frequently enough you can effectively lock the system into its initial state as it lacks the necessary amount of time to explore and evolve into other states. As you go into the extremities of short time intervals, the probability that the system will evolve into a different state approaches zero, and as such you have this freezing of the initial eigenstate. Take: |𝜓⟩ = ⍺|0⟩ + 𝛽|1⟩, say that we are measuring the system in the basis |0⟩, |1⟩ and imagine that we are to start in the state |0⟩. If measured frequently enough, you suffocate the system of the time required for evolution, as each measurement will keep collapsing it into the initial |0⟩ eigenstate and preventing it from ever transitioning into the |1⟩ state. Again, it is a bit counterintuitive since we typically associate the act of measurement as some sort of “interaction with the environment” synonymous with decoherence. Here the Zeno effect characterizes not the preservation of superposition, but a freezing of the initial eigenstate, whereby the system simply can’t evolve and thus is deprived of the opportunity to “decohere” in the traditional sense. Paradoxically, this rapid assault of frequent measurements does indeed collapse it into an eigenstate, but at least it is frozen and restrained from evolution such that you actually preserve the state rather than causing decoherence.


With this, we are now somewhat clued in to return to the pointer I had made earlier in regards to what Dr. Kalmijn wrote about positive feedback loop mechanisms as a means to excite/amplify weak electric signal, and tying it together with this breadcrumb of a suggestion that the Zeno effect alludes us towards. It is exceptionally beautiful to see how Kalmijn predicts the existence of these specialized excitable ion-channels long before the molecular tools had been available to confirm indeed the existence of these specially adapted voltage-gated potassium channels as we had discussed earlier, with their exceptional repolarization kinetics and resilience in supporting synaptic firing without exhaustion in diminished conductance. With hand-wavey insobriety, let us allow our imaginations to run free and unbounded from the commitment to too much mechanistic realism and the rigor of our no-go theorem prisons. To just enjoy the dance a little bit in dreaming up the loosest possible sentiment of how we might, as Poincaré bethought, *allow rough ideas to arise in crowds and collide with the glue of intuition for *it may be through science that we prove, but only through these playful acts of intuition that we discover. As with most of Biology, there are likely many components to this narrative with many overlaid functionalities to this one shared property, and as such let us not be too hard on ourselves in sketching a mechanism any less than picture perfect.

Picture this: a room full of 1000 divinely talented percussionists, each beating their drum at an arbitrary 150 bpm (the fastest drummer holds a record speed of 2730 bpm, so this should be an easy task. In fact most songs tick anywhere between 100–160 bpm, which not just by coincidence, is exactly correlative to that of the human heart- since that is exactly the neurological rhythm it targets). Again, from their supreme talent let us suppose that they are able to heed the directions of some madman conductor, who queues each of the 1000 drummers in to finely spaced intervals whereby no two drummers overlap at the same moment- evenly distributed across time. While the individual tempo of each drummer remains at 150 bpm, the collective (effective) beat frequency is now 150,000 bpm- assuming that they are able to maintain these perfectly staggered intervals. By recruiting a large collective of drummers– who, on their own, are already considered fast– with some cleverly precise orchestration, we have effectively constructed an ultra-high speed ensemble metronome. Bringing this back to the context of shark electroreception, we are now able to speculate in greater detail of what great mechanistic implication these specialized potassium channels may actually contribute to the spookiness of electroreception.

Hopefully the rhetoric about the talented drummers has helped sculpt some intuition for you in what functionality these potassium channels may yield, and how they ought to string the bow with the aforementioned quantum Zeno effect. As described earlier in the 2) bulletpoint about speed, the hallmark of quantum phenomena is not so much just the magnitude/size of signal but also the ultra-fast time domain in which it occurs, therefore we can surmise that any sensory faculty entwined to pick up on such phenomena must possess capabilities which resemble ultra-fast detectors. With that, we can imagine more vividly where and how to nest the aforementioned metronome effect into biophysical reality, that it effectively serves as an ultra-fast detector as well as a positive feed-back chamber to capture and amplify even the smallest of field disturbances. Just as with Zeno’s arrow paradox, one might imagine cinematically the radar display on some naval craft in entertaining the analogue of the potassium channel metronome as the detector’s “refresh rate”, whereby an apparatus with an extremely fast “refresh rate” would be able to discern even the slightest and briefest dynamics of some target object as opposed to one with a low “refresh rate” where they would observe the target object seemingly disappearing and reappearing across the screen upon every updated tick. This rapid firing potassium channel metronome in its collective beat frequency acts like that of an ultraspeed detector constantly ticking, with every voltage-dependent ion channel ready to pick up on even the slightest or most briefest changes of surrounding electric fields. One might speculate that these resiliently rapid-firing specialized synapses amassed into electroreceptive ensemble is like shooting with an ultra-high frame rate camera, of which– as mentioned in 2), if it can shoot at a femto-pico second frequency– could beat the rate of quantum decoherence and unveil all that is obscured to slower eyes in being able to track decoherence pathways. Even further, as Dr. Kalmijn so presciently alludes to, we can cast an even more ambitious picture not just of an ultra-fast detector, but also a signal preservation mechanism utilizing the quantum Zeno effect whereby some sort of positive feedback loop is accentuated by the rapid firing ion channels. Just as we envisioned the rapid ion channel firing as a means of high-speed detection, we can further imagine the persistent firings as generating domino-effect oscillations whereby one ion channel’s depolarization/repolarization triggers the voltage-dependent action of the other ion channels, leading to a cascading positive feedback loop of synapses firing en-masse in response to but a tiny stimulus. This positive feedback loop is essentially an amplification mechanism aiming to preserve– as the quantum Zeno effect does– some initially miniscule signal (in this case, some minute field disturbance). Just as the quantum Zeno effect aims to preserve an initial eigenstate by constant bombardment of measurements, one can imagine the ion channel metronome also as continuously detecting small field variations (akin to frequent measurements) as well as the ability to lock in a particular state and preserving it via feedback loop amplification, preventing it from fading away much the way the Zeno effect seeks to prevent quantum states from evolving away into decoherence. The dual complementarity features of these particularly excitable ion channels is that they both provide a means of readiness and rapid response– which can be seen as a biophysical analogue to the QZE, where frequent “measurement” or interaction with the signal stabilizes it– as well as the positive feedback amplification mechanism which ensures that the signal escalates to a level of neurological perception, allowing the organism to detect even the faintest of electric fields.
Anyways… Back to April 2nd 2023
Consider then, the unfathomably significantly more advanced system of olfaction in the animal kingdom. It is here that I come back to sharks. Colloquially, we are all at least somewhat aware of the substantially incredible sense of olfaction that sharks possess. “Hey did you know that sharks can smell blood from a mile away?”, but that conversation ends there in most typical scenarios. What particularly ignites my interest about sharks is that they possess yet another sensory system that is also barely understood. The Ampullae of Lorenzini (spotted pores on the snout of a shark, discovered by the late UCSD Scripps’ Adrianus Kalmijn) and the lateral line (loosely analogous to the “spinal cord” of a shark) seem to display abilities that absolutely break all comprehension and even physical law as we know it. The Ampullae of Lorenzini is responsible for the incredible acuity of electroreception (fields as small as 5nV/cm) present in sharks. This, in conjunction with the lateral line’s sensory capabilities in detecting the tiniest of harmonic disturbances in the vibration of water, makes sharks the incredible apex predators they are.
This does not make sense. According to physical law, such a perturbation in the electrodynamics and acoustics of such a far-away prey should not be able to be cohesively detected. It seems outside of the imagination that such a biological system can accurately decipher and preserve quantum decoherence from quantum coherence when considering especially that the stochastic noise-levels in the medium of the aquatic environment dwarf the intended signal (e.g: fish heartbeat) in amplitude by several magnitudes. There is a suggestion that there may be a quantum coherence occurring between the cryptochromes and/or chromophores inside the shark’s electroreceptors, in which they juggle various energy levels and spin-states of electrons via absorption of light (conserved mechanism in photosynthesis and cellular metabolism?), effectively amplifying the ability to detect weak electrical fields (Edit 2024: Interestingly, this suggests that the mechanism underlying Shark Olfaction, Electroreception, Magnetoreception may not necessarily be distinct faculties. Ultimately they are all representative of an underlying decoherence question. Poetically one might consider them analogues of the canonical double-slit, the Bohm-Aharanov, and the Stern-Gerlach experiment respectively).
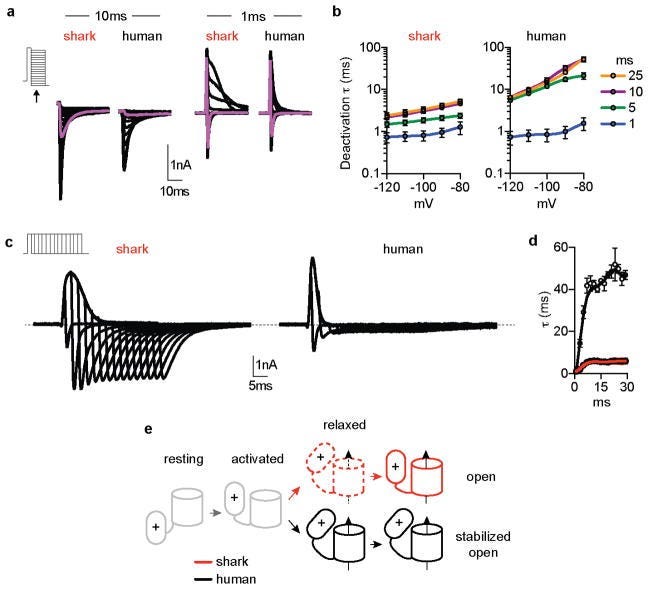
Sharks possess a uniquely different synaptic structure commonly referred to as “ribbon synapses”. There exists highly specialized and uniquely expressed potassium channels that differ in the canonical dynamics of almost all other species, in that they can be stimulated by quantitatively large and repetitive electrochemical voltage spikes. They display anomalously fast deactivation and activation kinetics, seemingly unperturbed and unexhausted by repeated firings shown by the potassium conductance failing to diminish even after substantial abuse of synaptic activation/deactivation (Edit 2024: In hindsight of the recent atto-second Nobel prize, and its femto-second predecessor, this fact seems to ring relevance. If the femto-second range is what defines electron, photon, interactions and QM can be considered ultimately to be a certain time-interval strata of ultrafast Physics, might the rapidly stacked action potentials of these specialized synapses provide some insight in how Sharks are able to map decoherence pathways?)
Even so — with all due respect to current Ichthyologists — this still does not quite make sense when you consider the nullifying effects (since dielectric mediums actually further accentuate electric fields, although not necessarily for the benefit of coherency) of transferring/detecting signals in a charged dielectric medium like seawater, combined with the vast distances seeming to ignore entirely the inverse square law. The Shark’s ability to detect electric fields akin to two AA batteries connected 1000 miles apart is in some sense the BioPhysical analogue of the Bohm-Aharanov experiment: the fascinating experiment revealing that the vector & scalar potential fields of Maxwell’s electromagnetism are not merely Mathematical aids but are in fact real Physical structures. The existing scientific literature has certainly identified a few of these specialized shark potassium channels, but even so they conclude that there must exist an abundance of missing auxiliary protein subunits, signaling molecule cascades, or other functional biochemical components, in order to cohesively support and construct a plausible regulatory mechanism of shark electroreception. It seems most definitely certain to me that the sensory faculties equipped by sharks must utilize some (or many) strange unknown physical principles that appear to bypass our current understanding of Physics.
In Layman’s terms: the underwater world is full of noisy, random, incoherently obtrusive sounds and electrical signals. How is it possible that a shark can hear/smell/feel these signals despite being drowned out in a whole rock concert of stimulatory events?
It is at this point that the sufficiently attentive biologist may say: “This is all true Neil, but what makes sharks so special? Why not simply study the olfactory/sensory systems in drosophila or laboratory mammals?” My answer to that is simple: evolutionarily conserved mechanisms should be investigated in species of varying phylogenetic age. It is a largely overlooked fact that sharks as a species are incredibly phylogenetically old. Trees as a species are considered to have entered the earthly scene around 350–380 million years ago. Sharks on the other hand, first began swimming the oceans roughly 450 million years ago. The geneticist reading this will immediately understand the significance of the temporal relic of an organism that is the shark. Sharks can be considered to be potentially one of our eldest ancestors on the evolutionary divergence tree. It seems agreeably clear to me that the biochemical facilities that evolution developed in sharks have turned out to be immensely successful, considering that they are still swimming our oceans today. It is also no longer a disputed notion that nature herself is the master of utilizing quantum phenomena to her advantage, and from this I draw the possibility of making the case that sharks may very well be the appropriate model organism to study in regards to defeating the quantum roadblock.
For very long I had placed my hopes and bets on the shoulders of those working in mathematics and particle/theoretical physics to remove the quantum roadblock and allow us scientists of other sectors to continue investigating the true nature of this odd reality. While I respect most honorably the efforts of people like Prof. Lee Smolin, Dr. Stephen Wolfram, Dr. Garret Lisi, etc. in their attempts to bridge the gap between Einstein and Bohr, I personally and humbly resign on simply waiting for somebody else to build me the tools to gather information and harvest the truth. My new take on this is inspired by that of Dr Brian Keating, a cosmological experimentalist at UCSD who presents the argument along the lines of: “Why build a gigantic expensive hadron collider? Mother nature herself provides us with the best laboratory and high-energy particle physics experiments in the cosmos of space”. Perhaps also inspired by the quantum magnetic spin-sensitive navigation systems in pigeon brains (Pigeons are basically performing the Stern-Gerlach experiment in-vivo) and the immunological antibody design breakthroughs made through snake venom, I see now that mother nature has provided the curiosities of the quantum biologist with a vast data set of potential discovery and experiment in the remarkable kingdom of flora and fauna. Instead of waiting for fundamental theories to explain our natural phenomena, perhaps we must investigate natural phenomena in order to find said fundamental theories.
There are not many people in this world who share my passionate interest for both molecular magic and sharks, but even so I can’t say that there’s any better place to investigate this than at UCSD, who leads the world in both the marine sciences and the molecular sciences. I am aware that this is quite the long-shot, trust me, my self-doubts are no quiet critics. There is the very large possibility that I am extraordinarily incorrect about everything I have said, but even so I think that in the letter and spirit of science we should not be afraid to put forward ideas that may contain errors, for now that this escapes into the world it cannot be taken back. I welcome the critics who may present newfound information that implies the vindication or extermination of my case, but nevertheless this shall remain on the public domain in the event that a recovery is needed.
Many times I have been asked by friends and family what exactly it is that I intend to do with my biochemistry career, where I’ve given all sorts of answers ranging from medicine, biotech, pharmaceuticals, research, or sometimes even the plain “I don’t know but we’ll see”. It has always been said that the potential applications and integral relevances of biochemistry are far and wide, which served for me as an effective mask to avoid thinking about what it is that actually stimulates my desire for this introspectively personal conquest. The honest truth is, I have no real intentions on what to do with this. The idea of slaving away in industry pharma spending my time designing synthesis routes for useless modifications to a drug in order for hefty profitable patents to be made for NASDAQ listed corporations, never really sat right with me. My honest confession is that I am but a child who simply enjoys reading and exhausting his own curiosities.
Cicero(?)’s Dubitando ad veritatem pervenimus (“It is by way of doubting that we arrive at the truth”) is the hallmark quote of every heart-filled scientific investigator. To the aspiring investigator who reads this, I implore you not to seek that I am right or wrong, but to live honorably in finding the truth. I wish you the best of luck.
Neil Tendian
04/02/2023